









«Насколько я знаю, все физические результаты
a priori имеют свои источники в симметрии»
Герман Вейль
«Развитие Вселенной с момента ее возникновения выглядит
как непрерывная последовательность нарушений симметрии…
Феномен жизни естественно вписывается в эту картину».
Фриман Дж.Дайсон

Выдающиеся достижения биохимии и молекулярной биологии последних 50–70 лет в изучении жизненных процессов заслонили принципиальные для биологии фундаментальные физические принципы организации и функционирования живых систем, сформулированные в 30-е – 60-е годы прошлого столетия. Каким-то образом исчезло из рассмотрения важнейшее для биологической эволюции положение: естественный отбор в биологии происходит по физическим, а не по химическим критериям. И самые общие принципы построения живого — дискретность, неравновесность, нелинейность, синергизм, иерархичность, машинность, сочетание фазовых состояний, а также квантовых и макроскопических масштабов и процессов, системность и пр. — являются физическими. В свою очередь, универсальный «химический» инструментарий позволяет эффективно реализовать известные на Земле «биологические» формы жизни.
Идеи симметрии в физике играют фундаментальную роль при рассмотрении природных закономерностей на самых разных уровнях организации материи. Столь же естественно считать представления о симметрии и нарушениях симметрии фундаментальным фактором при рассмотрении физических основ строения живых систем, их происхождения и эволюции. Симметрические характеристики упорядоченности системы на одном её уровне предопределят её масштаб и симметрические характеристики на следующем иерархическом уровне эволюционного развития.
Особое место в категории симметрий-асимметрий занимает понятие хиральности. В нашем рассмотрении молекулярно-биологических систем мы будем пользоваться классическим определением хиральности: хиральность — свойство молекулы или объекта быть несовместимым со своим зеркальным отображением при любых комбинациях перемещений и вращений в трёхмерном пространстве. В энантиомере (энантиоморфе) отсутствует симметрия относительно сторон и осевая симметрия.
Хиральность мономеров, составляющих основу информационно-детерминированных биополимеров, белков и нуклеиновых кислот, обычно воспринимается как иррациональный эпизод в целесообразном эволюционном процессе, как «плата» за уникальную способность углерода образовывать огромное количество неорганических и органических соединений, гармонично связывающих в единое целое структуры и функции живых систем. А гомохиральность первичных структур воспринимается как тривиальный выход из дуалистической ситуации. Так, известно, что гомохиральность входящих в белки «левых» L-аминокислот обеспечивает стереоспецифичность комплементарных взаимодействий и минимизирует объём информации, необходимый для однозначного кодирования нуклеиновыми кислотами аминокислотных последовательностей. Также общеизвестно, что дезоксирибоза и рибоза, входящие в состав уникальных ДНК и РНК, являются «правыми» D-изомерами.
Мы выдвигаем принципиально новую точку зрения на проблему хиральности: хиральный дуализм является фундаментальной основой структурообразования в молекулярной биологии, а хиральность названных соединений — аминокислот, рибозы и дезоксирибозы — есть важнейший инструмент иерархической стратификации внутримолекулярных и надмолекулярных структурных уровней, а также специфичности межмолекулярных взаимодействий.
В макромолекулярных системах нуклеиновых кислот и белков нами впервые были выделены как целое, как хиральные инварианты молекулярной биологии знакопеременные иерархии хиральных структур в последовательностях от «нижнего» ассиметричного атома углерода в sp3-гибридизации до суперспиралей и надмолекулярных структур. Отмечено чередование знака хиральности D-L-D-L при переходе на более высокий уровень структурно-функциональной организации ДНК. Две базовые первичные полимерные цепочки ДНК составлены молекулами D-дезоксирибозы. Цепочки соединены нуклеиновыми основаниями в «левой» гош-конформации, а все вместе они образуют известную «правую» двойную спираль ДНК. У бактерий кольцевая «правая» спираль скручивается в «левую» суперспираль.
У белков первичная цепочка аминокислот — «левая». Образуя вторичные структуры, они скручиваются в «правые» α-спирали, которые, в свою очередь, сворачиваются в букеты «левых» суперспиралей. Четвертичные структуры белков, собирающиеся уже из самостоятельных молекул, снова образуют «правые» витые конструкции. У белков последовательность смены знака хиральности в структурно-функциональной иерархии: L-D-L-D.
Посмотрев на рисунок, мы можем убедиться, что центральный блок всей молекулярной биологии, представленный знакопеременными хиральными иерархиями внутри- и надмолекулярных структур ДНК и белков, образует периодическую таблицу. Вследствие смещения столбцов на пол-периода, это «ядро» молекулярной биологии обретает ось симметрии и становится ахиральным блоком. Таким образом, молекулярная биология всех земных форм жизни является симметрически замкнутой знакопеременной хиральной периодической системой.
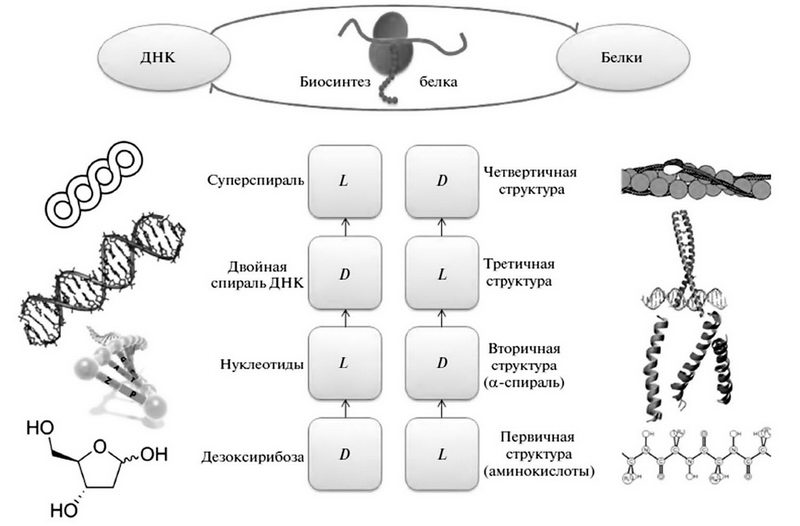
Периодическая таблица знакопеременных иерархий хиральных (спиральных) структур от первичной до четвертичной для ДНК (левая колонка) и белков (правая колонка): L — левая конфигурация энантиомера или спирали, D — правая.
Формирование знакопеременных хиральных иерархий в макромолекулярных структурах обусловлено четкой физической причиной — стремлением системы понизить исходный уровень свободной энергии, образующийся при энергозависимом отборе гомохиральных мономеров первичных структур из их рацемических смесей. Заметим, что в неживой природе имеется множество примеров спонтанного формирования подобных иерархий и что живая природа успешно акцептировала этот принцип.
Знакопеременная хиральная иерархичность сопряженных уровней структур в макромолекулах имеет общебиологическую значимость: обуславливает дискретность этих уровней, служит инструментом пространственной самоорганизации (уникальной самосборки), структурной основой «выделенных механических» степеней свободы в конструкциях биологических макромолекулярных машин.
Литература
1. Твердислов В.А. Хиральность как первичный переключатель иерархических уровней в молекулярно-биологических системах / Биофизика. 2013. Т. 58, No 1. С. 159–164
2. В.А.Твердислов, А.Э.Сидорова, Л.В.Яковенко От симметрий — к законам эволюции.1. Хиральность как инструмент стратификации активных сред. Биофизика, 2012, том 57, вып. 1, С. 146 — 154
3. Твердислов В.А., Малышко Е.В., Ильченко С.А. Известия РАН. Серия физическая, 79 (3), 2015, 1728
В.А.Твердислов,
зав. кафедрой биофизики